Chapters
Lodish 4th edition: Chapter 21
Moyes and Schulte: Chapter 6 pages 207 - 245
Moyes and Shulte: Chapter 9 pages 362-368
Muscle cells
This course will discuss the three different muscle types:
1. Skeletal muscle
2. Cardiac muscle
3. Smooth muscle

Figure 22-40, Molecular Biology of the Cell, 4th edition. The four classes of muscle cells of a mammal.
(A) Schematic drawings (to scale).
(B-E) Scanning electron micrographs, showing (B) skeletal muscle from the neck of a hamster, (C) heart muscle from a rat, (D) smooth muscle from the urinary bladder of a guinea pig, and (E) myoepithelial cells in a secretory alveolus from a lactating rat mammary gland. The arrows in (C) point to intercalated discs—end-to-end junctions between the heart muscle cells; skeletal muscle cells in long muscles are joined end to end in a similar way. Note that the smooth muscle is shown at a lower magnification than the others.
Vertebrates and many invertebrates have three main classes of muscle — skeletal, cardiac and smooth. Skeletal muscles connect the bones in the arms, legs, and spine and are used in complex coordinated activities, such as walking or positioning of the head; they generate rapid movements by contracting quickly. In this type of contraction, termed isotonic contraction, the muscle shortens as force is generated. Another major function of skeletal muscle is to hold objects immobile. In the clenching of fists or tensing of muscles, for example, pairs of contracting muscles work to oppose each other and thus cancel out any movements. In such isometric contraction, the overall length of a muscle remains constant but its tension increases. Smooth muscles surround internal organs such as the large and small intestines, the uterus, and large blood vessels. The contraction and relaxation of smooth muscles controls the diameter of blood vessels and also propels food along the gastrointestinal tract. Compared with skeletal muscles, smooth muscle cells contract and relax slowly, and they can create and maintain tension for long periods of time.
Skeletal Muscle Contraction
A general overview of the membrane potential changes that occur in the
skeletal muscle fiber out lined below. Each part will be discussed in greater
detail in the following sections.
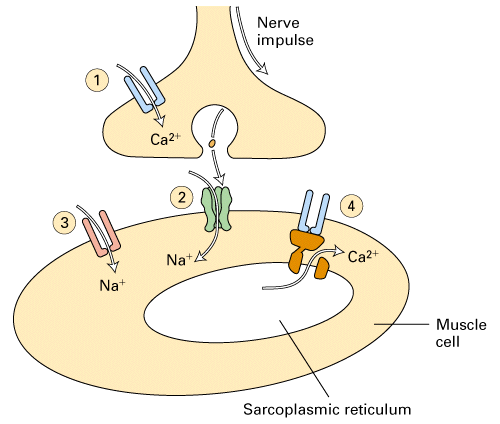
Figure 21-37, Lodish 4th edition. Activation of gated ion channels at a neuromuscular junction.
1) Arrival of the action potential at the presynpatic side of the synapse
opens voltage gated Ca+2 channels and triggers vesicle fusions and thus
acetylcholine release. See section on NMJ.
2) Acetylcholine binds to its ligand-gated ion channel which opens to allow
Na+ to flow into the muscle cell. See section on acetylcholine
receptor.
3) The depolarization from the entry of Na+ into the muscle if it reaches
threshold will open the voltage-gated Na+ channels in the muscle cell. See
section on muscle channels and action potentials.
4) The action potential generated by the muscle voltage-gated Na+ channels
spread throughout the muscle cell and opens voltage-gated Ca+2 channels to
allow Ca+2 to flow from the sarcoplasmic reticulum into the cytosol. The rise
in cytoplasmic Ca+2 causes muscle contraction through actin and myosin. See
section on actin, myosin and sliding filament model.
Skeletal muscle (striated muscle)
Skeletal muscle cells are one of the largest cells in the body. They are multinucleate formed by the fusion of myoblasts. Diameters range from 50 to 150 microns with lengths ranging from mm to cm.
To be effective the whole muscle fiber develops tension at the same time
and of course to generate force the muscle fiber is anchored.
Muscle fibers contract in response to an electrical signal, depolarization. The
signal is generated at the synapse (the neuromuscular junction) and propagated
through an action potential via the muscle fiber membrane. The membrane of the
cell has specialized invaginations called Transverse-tubules (T-tubules) that
enter into the cell (at every 1-2 microns). Thus the action potential can be
rapidly transmitted deep into the interior of the cell resulting a delay of
only 3-5 msec between the depolarization at the synapse and the first muscle
fiber tension. The T-tubule network is so extensive that 50-80% of the plasma
membrane is in the T-tubules.

Figure 18-31. The sarcoplasmic reticulum (SR) regulates the cytosolic Ca2+ level in skeletal muscle.
(a) Three-dimensional drawing of a portion of a muscle cell (myofiber) composed of six myofibrils. The transverse (T) tubules, which are invaginations of the plasma membrane, enter myofibers at the Z disks, where they come in close contact with the terminal cisternae of the SR, forming triads. The terminal cisternae store Ca2+ ions and connect with the lacelike network of SR tubules that overlie the A band. (b) Release and recovery of Ca2+ ions by the SR. Depolarization of a muscle cell (step 1) induces the release of Ca2+ ions stored in the SR via Ca2+ release proteins in the SR membrane (step 2). Subsequently, Ca2+ ATPases in the SR membrane pump Ca2+ ions from the cytosol back into the SR, restoring the cytosolic Ca2+ concentration to its resting level within about 30 milliseconds (step 3).
The plasma membrane containing the ion channels sends invaginations (called
T tubules) into the myofibers at the point of the Z disks. The T tubule
contacts the sarcoplasmic reticulum forming a structure called a triad. At
this point in the plasma member is a voltage-gated Ca+2 channel (dihydropyridine
receptor) which is associated with the Ca+2 release channel (ryanodine
receptor) in the sarcoplamic recticulum membrane.
Vertebrate skeletal muscle is striated given the arrangement of action and
myosin in each sacromere.
We will discuss at length the function of the muscle fiber from the generation
of the depolarizing endplate potential to the triggering of the myosin/actin
interactions to generate contraction.
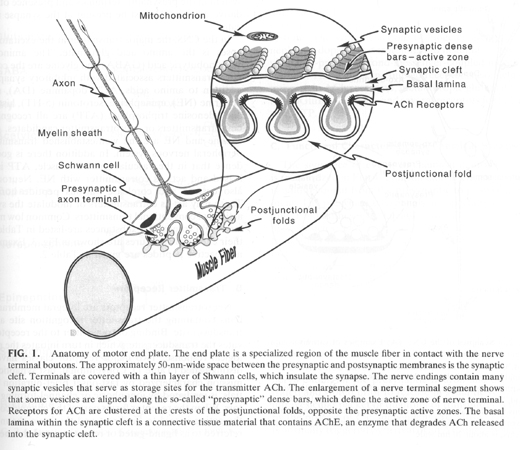
Neuromuscular junction
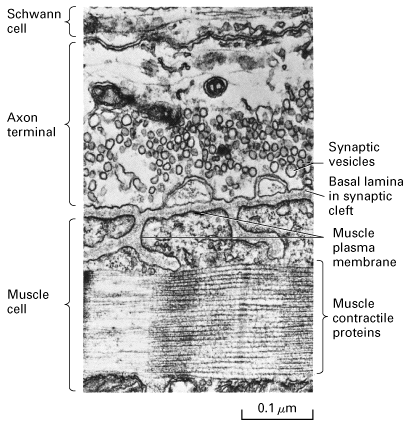
Figure 21-33, Lodish 4th edition. High magnification image from a frog neuromuscular junction.
From "Cell Physiology Source Book"
Each muscle fiber is innervated by a motorneuron. One motorneuron can innervate one or multiple fibers. Each motorneuron plus its complement of muscle fibers is called a motor unit as well all contract in concert.
The synpase between the motorneuron and the muscle fiber is called a neuromuscular junction (NMJ) and is a unique type of synpase. The motor nerve terminal branches run in shallow grooves on surface of muscle. The nerve terminal contains many mitochondria and vesicles which can be seen lined up in double rows along side the voltage-gated Ca+ channels attached to presynaptic membrane => active zone.
The vesicles of the NMJ have very high concentrations of neurotrasmitter
(2,000 to 10,000 molecules of acetylcholine per vesicle). The function of the
NMJ is to generate a contraction in the muscle fiber. There an excess of
neurotransmitter is released to ensure that the resulting post-synaptic
depolarization is strong enough to generate an action potential - "safety
margin".
The average post-synaptic potential in a muscle fiber is a depolarization of
40 mV. The post-synaptic depolarization is always much greater than threshold
for an action potential (rest is -80 to -70 mV and threshold for an action
potential is -60 to -50 mV on average).
The post-synaptic membrane of the muscle has folds which are full of the
neurotransmitter receptor which is:
acetylcholine is the neurotransmitter for the mammalian NMJ
glutamate is the neurotransmitter for the insect NMJ
Acetylcholine receptor
Studies to look at the numbers of acetylcholine receptors found ~15,000
receptors per micron2. To do these studies researchers used a toxin
that is high specific for Ach receptors called alpha-bungarotoxin. Alpha-bungarotoxin
is another example of a snake venom used by the predator to kill its prey.
Researchers use the toxin by labeling the toxin with either 125I
or a fluorescent label, applying the toxin to a NMJ preparation and then
assaying where the receptors are by following the radio label or the
fluorescence label.
These types of studies showed a very high concentration of receptors in the
synapse but a complete absence of receptors away from the synapse.
The molecular nature of the acetylcholine receptor is well studied as you
might except these proteins are highly conserved between all animals.
The acetylcholine receptor is made up of five subunits (all have been cloned):
2 alpha, 1 beta, 1 gamma and 1 delta subunit.
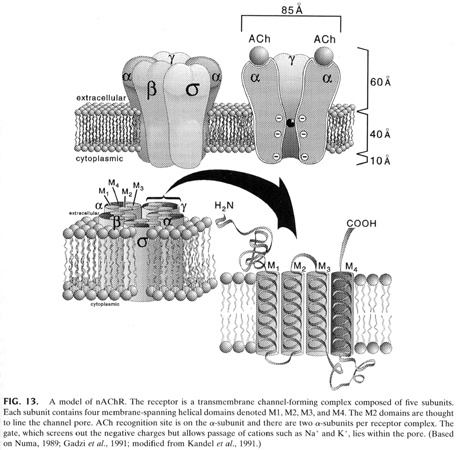
From "Cell Physiology Source Book"
The receptor is made up of 4 different transmembrane proteins one of which
(the alpha subunit) is repeated to give 5 subunits to create the ion channel.
Acetylcholine binds to the alpha subunit and thus it takes two molecules of
acetylcholine to open the channel.
One acetylcholine receptor opens and allow 1.5 x 104 Na+ ions/msec
of open time. The channel opens on average 1 msec. Therefore one open channel
causes a depolarization of about 0.3 mV. From other
studies it has been shown that the amount of neurotransmitter contained in one
vesicle causes an post-synaptic potential of ~ 1 mV. Therefore 1 vesicle
contains enough neurotransmitter to open ~3000 receptors and because two
molecules of Ach is needed to open one receptor there must be a minimum of
~6,000 molecules Ach per vesicle.
If the average depolarization generated at a NMJ of a muscle fiber is 40 mV
then there must be at least 40 vesicles released and in the order of 120,000
receptors activated at the NMJ.
From "Cell Physiology Source Book"
The postsynaptic response of the muscle fiber is the same as an epsp in a neuron except that it is called an endplate potential (epp) or an excitatory junctional potential (ejp). The influx of Na+ through the receptor causes a depolarization that is normally sufficient to open the voltage-gated Na+ channels found in the muscle fiber.
To record and analyze the endplate potential researchers first have to remove the influence of the voltage-gated Na+ channels which of course generate action potentials that would mask any of the smaller membrane potentials made by the acetylcholine receptors. To do this researchers simply add TTX which is a highly specific toxin that blocks voltage-gated Na+ channels. Now in the absence of action potentials the membrane potentials generated by the acetylcholine receptor can be measured.
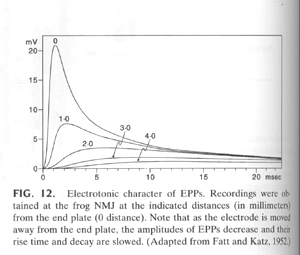
The above figure shows the recorded endplate potentials from a muscle fiber with a recording electrode at the NMJ (0) and 1, 2, 3 or 4 mm away. The amplitude of the epp is reduced the further away from the initial depolarization. This is precisely the same mechanism that we saw in the dendrites of neurons and the same sources of current loss (i.e. cable properties, Cm, Rm and Ri) are in effect here. The length constant can be determined from these types of experiments and on average the length constant is ~ 1 mm.
Remember though the muscle can be mm to cm in length and needs to contract rapidly and smoothly i.e. across its entire surface not just in one spot. Thus the presence of voltage-gated Na+ channels are essential to propagate the signal rapidly without loss of signal to the entire muscle surface. The presence of the T-tubules delivers the action potential deep into the muscle interior.
Finally the NMJ is usually centered in the middle of the muscle fiber resulting in action potentials following in two directions away from the NMJ.
Striated muscle channels and action potentials
From "Cell Physiology Source Book"
The skeletal muscle has a similar array of voltage-gated ion channels, transporters and pumps as the neurons we discussed in previous sections.
Pumps and transporters - these
were discussed in the previous lectures. In particular the muscle fiber has:
1) Na+/K+ ATPase pump - to establish the electrochemical gradients of Na+ and
K+
2) Ca+2 ATPase pump - uses energy from ATP to remove 2 Ca+2 from the inside to
the outside of the cell to ensure that internal Ca+2 concentrations remain low
(10-7 mM internal).
3) Na+/Ca+2 cotransporter - to also remove Ca+2 from the inside of the cell
and uses the energy from the cotransport of 3 Na+ molecules to export 1 Ca+2.
4) Muscle Ca+2 ATPase pump - a different pump from number 2 above. Found
highly concentrated on the sarcoplasmic reticulum (SR) (constitutes 80% of the
protein found in the SR membranes). The muscle Ca+2 ATPase pumps 2 Ca+2 into
the SR to lower cytosolic Ca+2 and to concentrate Ca+2 into internal stores.
Channels - muscle cells share
many of the same ion channels as neurons.
1) leak channels - besides the leak K+ channel, skeletal muscle cells have a
high concentration of Cl- leak channels so much so that the resting membrane
potential is usually the same as the Nernst potential for Cl- (around -80 mV
in these cells). The high permeability to Cl- helps repolarize the membrane
after an action potential.
2) voltage-gated Na+ channels - there is a skeletal muscle voltage-gated Na+
channel which properties very much like the neuronal voltage-gated Na+ channel
we have already discussed at length. TTX - sensitive as we saw above. This
channel is of course responsible for the production of the action potential.
Once opened the influx of Na+ through the channel further depolarizes the
membrane thus opening more Na+ channels and this regenerative cycle continues
until the Na+ channels start to inactivate and the delayed K+ channel begins
to open.
3) voltage-gated K+ channel - the delayed rectifier K+ channel which we have
already seen in neurons with exactly the same properties. In other words has a
high threshold and needs a strong depolarization to open and works to bring
the membrane back to resting potentials.
4) voltage gated Ca+2 channels - high threshold Ca+2 channels very much like
we saw before at the neuronal chemical synapse. Needs a strong depolarization
to open and very slowly inactivates. These Ca+2 channels are sensitive to DHP
(dihydropyridine) and are found concentrated in the T-tubules.
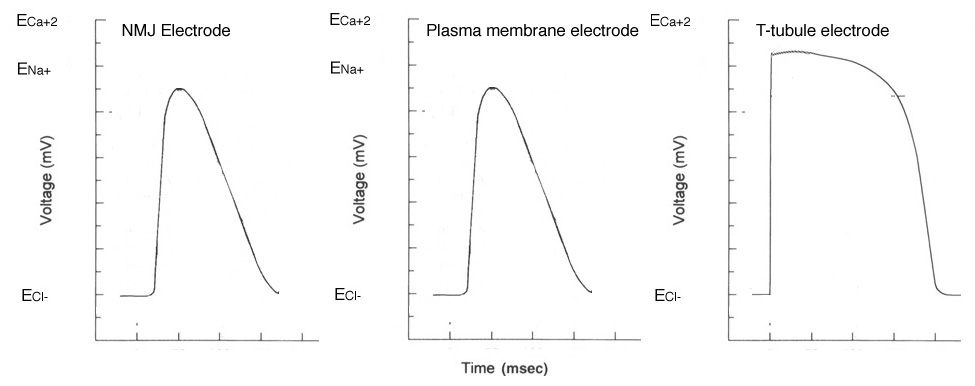
Action potentials recorded at the NMJ, the muscle plasma membrane and in the T-tubule
From "Cell Physiology Source Book"
The average post-synaptic potential in a muscle fiber is a depolarization
of 40 mV. The post-synaptic depolarization is always much greater than
threshold for an action potential (rest is -80 to -70 mV and threshold for an
action potential is -60 to -50 mV on average). This is to ensure that an
action potential will be generated in the muscle fiber when ever a signal
arrives at the NMJ.
Triads and Sarcoplasmic reticulum
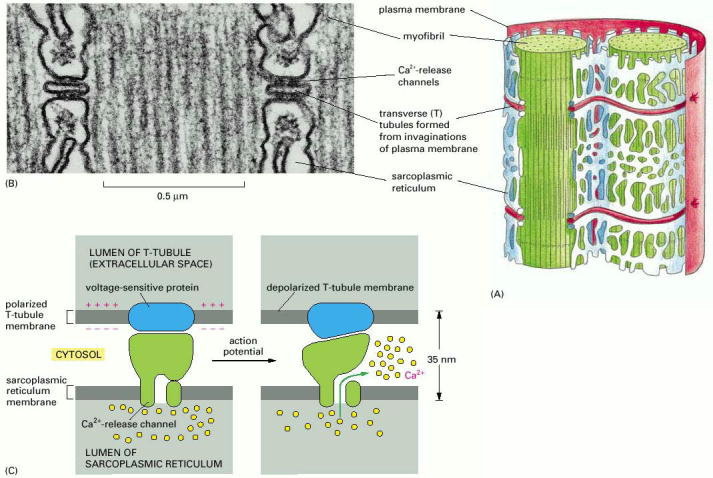
Figure 16-73, Molecular Biology of the Cell, 4th edition. T tubules and the sarcoplasmic reticulum.
(A) Drawing of the two membrane systems that relay the signal to contract from the muscle cell plasma membrane to all of the myofibrils in the cell.
(B) Electron micrograph showing two T tubules. Note the position of the large Ca2+-release channels in the sarcoplasmic reticulum membrane; they look like square-shaped “feet” that connect to the adjacent T-tubule membrane.
(C) Schematic diagram showing how a Ca2+-release channel in the sarcoplasmic reticulum membrane is thought to be opened by a voltage-sensitive transmembrane protein in the adjacent T-tubule membrane.
The link between depolarization and Ca+2 release or excitation-contraction coupling occurs at the junctions between the T-tubule and the sarcoplasmic reticulum. 80% of the T-tubules membrane is associated with the sarcoplasmic reticulum at triads. The voltage-gated Ca+2 channels are concentrated in the T-tubules in the triads. The Ca+2 release channel found in the sarcoplasmic reticulum membrane is associated with the voltage-gated Ca+ 2 channel at this point. This close association allows for the rapid signaling from action potential to Ca+2 release.
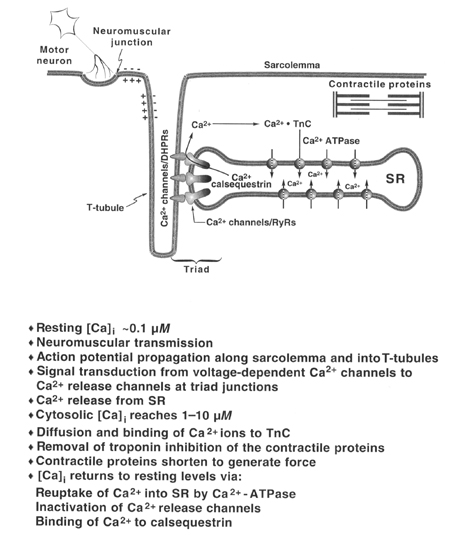
From "Cell Physiology Source Book"
The overall effect of an action potential arriving at the muscle fiber is that the voltage-gated Na+ channels in the membrane distribute the depolarization to the T-tubules which in turn depolarizes to open the voltage-gated Ca+2 channels. This results in a release of Ca+2 through the Ca+2 release channels from the sarcoplasmic reticulum. After the action potential is passed and the voltage-gated Ca+2 channels close, the Ca+2 release channels close and Ca+2 is recycled back into the sarcoplasmic reticulum through the Ca+2 ATPases.

Figure 20-40, Lodish 4th edition. Release of Ca2+ stores mediated by ryanodine receptors (RYRs) in skeletal muscle.
Voltage sensing dihydropyridine receptors in the plasma membrane contact ryanodine receptors located in the membrane of the sarcoplasmic reticulum. In response to a change in voltage, the dihydropyridine receptors undergo a conformational change; this produces a conformational change in the associated RYRs, opening them so that Ca2+ ions can exit into the cytosol.
The voltage-gated Ca+2 channel is either closely localized to or makes a
physical connection to the Ca+2 release channels in the sarcoplasmic
reticulum.
The voltage-gated Ca+ 2 channel is also known as the DHP receptor and the Ca+
2 release channel is also known as the ryanodine receptor. Both these
chemicals bind to their respective channels.
Not all Ca+2 release channels are associated with voltage-gated Ca+2 channels
and these non-associated channels are thought to be opened solely by Ca+2
influx into the cytosol from the voltage-gated Ca+2 channels.
The Ca+2 release channel in the sarcoplasmic reticulum of most muscle cells
(smooth, cardiac, skeletal) is a Ca+2 activated Ca+2 channel. In other words
the Ca+2 that enters through the voltage-gated Ca+2 channel from the outside
of the cell triggers the opening of the Ca+2 release channel resulting in an
efflux of Ca+2 from the sarcoplasmic reticulum into the cytoplasm.
What is interesting is that the Ca+2 release channel is stimulated to open at
with low concentrations of Ca+2 ( up to 0.1 mM) in the cytosol but inhibited
by high concentrations of Ca+2 in the cytosol (0.5 mM and higher for cardiac
cells). This means that as Ca+2 is released from the sarcoplasmic reticulum it
starts to inhibit the Ca+2 release channel. This is necessary otherwise the
Ca+2 release would be continual.
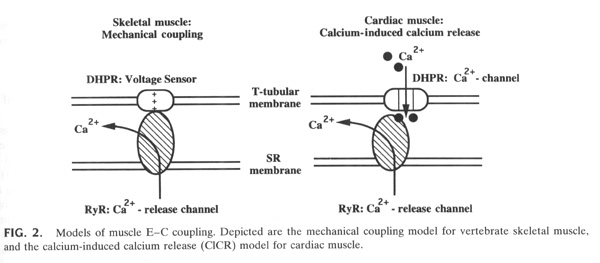
From "Cell Physiology Source Book"
In vertebrate skeletal muscle the Ca+2 release channel is stimulated to open in two ways through two different Ca+2 release channels: Ca+2 gated (see above in case of non-associated release channels) and mechanically gated. In vertebrate skeletal muscle cells the skeletal voltage-gated Ca+2 channel is physically linked to the Ca+2 release channel. As the voltage-gated Ca+2 channel goes through a protein conformational change during depolarization i.e as the voltage-sensor moves in the membrane during the depolarization, this opens both the voltage-gated Ca+2 channel and somehow this physical change is transduced to the the Ca+2 release channel.
Experiments:
The following are examples of experiments that were carried out to prove
the above models of muscle cell function.
These experiments are based on a simple assay system that tests muscle
function (i.e. contraction) under different conditions.
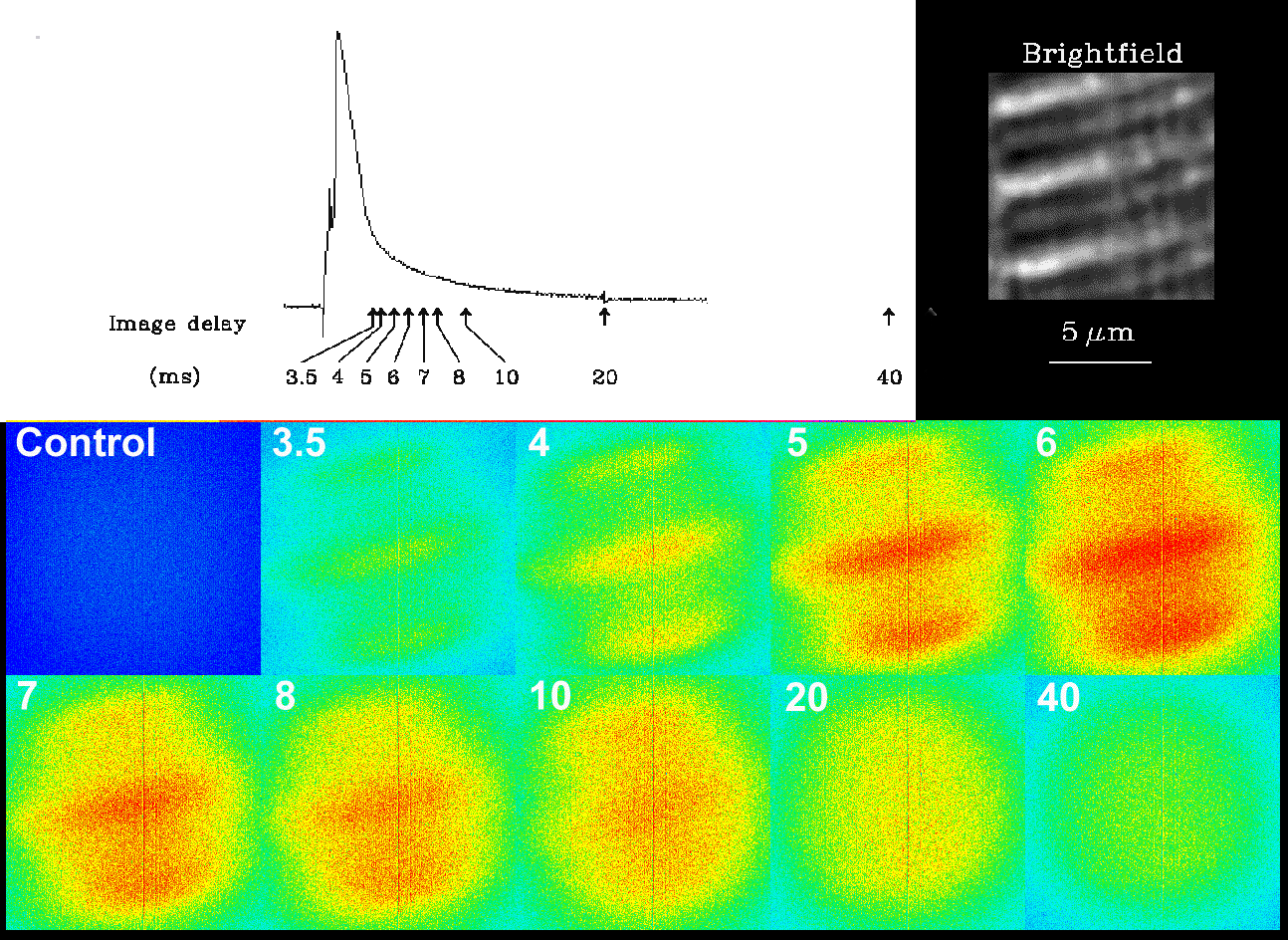
From Escobar et al. (1994) Nature 367, 739-741. Calcium gradients were measured by pulsed laser imaging.
Single skeletal muscle fibers from the semitendinosus muscle of the bullfrog Rana catesbeiana were mounted in an inverted double Vaseline gap chamber and held at -90 mV in current clamp. Depolarization induced an action potential (top left). The bottom panel shows a sequence of pseudocolored fluorescence images from rhod-2 loaded fibers taken after delays of 3.5, 4,5,6,7,8, 10, 20,25 and 40 ms, respectively (marked by arrows on the action potential trace). The brightfield image (top right) shows the sarcomere structure. Note that the gradient reflects calcium release from the terminal sarcoplasmic reticulum adjacent to the t-tubules.
From Escobar et al. (1994) Nature 367, 739-741. An animated calcium transient in a frog skeletal muscle fiber measured using pulsed laser imaging, showing the appearance and disappearance of intrasarcomeric calcium gradients.
To test the physiology of muscle fibers the muscle fiber is dissected out and attached usually via its tendon to a solid surface. At the other end a the force of contraction is measured. The muscle fiber can be stimulated to contract through a number of experimental means, either by stimulating the motorneuron that is synapsed with the fiber or by directly depolarizing the muscle fiber with an injection electrode (B). In either case the force of contraction can be measured.
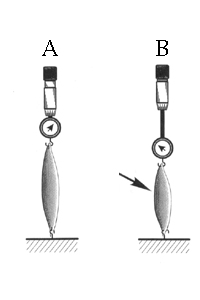
From "Cell Physiology Source Book"
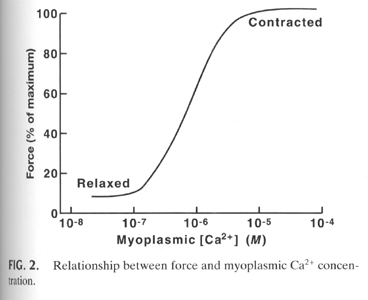
The degree of contraction or force exerted by the muscle fiber is dependent on the concentration of Ca+2 inside the muscle cell. As the Ca+2 concentration increases the force of the contraction increases to a maximal level.
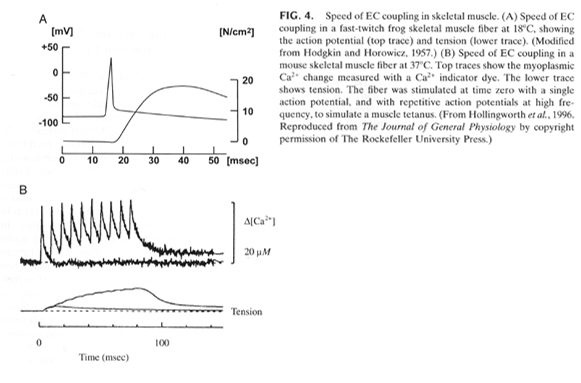
From "Cell Physiology Source Book"
The above figure highlights the timing between excitation-contraction
coupling. The first A shows the speed of the excitation (action potential) -
contraction coupling. Tension or contraction is generated within
milliseconds of the generation of the action potential.
In B shows the time scale of the Ca+2 peak in response to each action
potential. The fiber was stimulated with multiple action potentials. The
peak in Ca+2 release that occurs with each action potential was measured
using a Ca+2 sensitive dye such as Fura-2
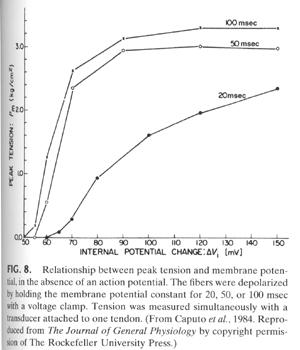
From "Cell Physiology Source Book"
Figure 8 details the experiments that show that (in the absence of external Ca+2) simply depolarizing the muscle cell membrane beyond Ca+2 channel threshold is enough to generate muscle contraction or force in a vertebrate skeletal muscle fiber. If this experiment was carried out in a cardiac muscle cell then there would be no force generated. Cardiac, smooth and invertebrate skeletal muscle cells depend on the influx of Ca+2 into the cell from the outside to trigger the opening of the Ca+2 release channel.
The above experiment in Figure 8 is one of the experiments that lead to the idea that it in vertebrate skeletal muscle cells it is not only Ca+2 entering through the voltage-gated Ca+2 channel that opens the Ca+2 release channel but must be a physical contact between the opening of the voltage-gated Ca+2 channel and the release channel.
Actin and myosin in muscle contraction
Release of Ca+2 triggers the contraction - provides the basis for
excitation-contraction coupling

Figure 16-69, Lodish 4th edition. Skeletal muscle myofibrils.
(A) Low-magnification electron micrograph of a longitudinal section through a skeletal muscle cell of a rabbit, showing the regular pattern of cross-striations. The cell contains many myofibrils aligned in parallel. (B) Detail of the skeletal muscle shown in (A), showing portions of two adjacent myofibrils and the definition of a sarcomere. (C) Schematic diagram of a single sarcomere, showing the origin of the dark and light bands seen in the electron micrographs. The Z discs, at each end of the sarcomere, are attachment sites for the plus ends of actin filaments (thin filaments); the M line, or midline, is the location of proteins that link adjacent myosin II filaments (thick filaments) to one another. The dark bands, which mark the location of the thick filaments, are sometimes called A bands because they appear anisotropic in polarized light (that is, their refractive index changes with the plane of polarization). The light bands, which contain only thin filaments and therefore have a lower density of protein, are relatively isotropic in polarized light and are sometimes called I bands.
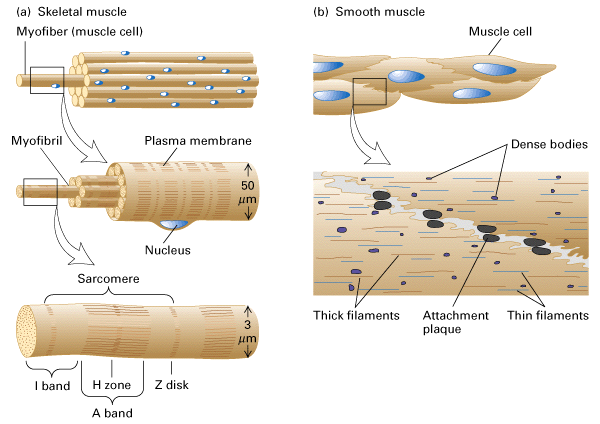
Figure 18-26, Lodish 4th edition. General structure of skeletal and smooth muscle.
(a) Skeletal muscle tissue is composed of bundles of multinucleated muscle cells, or myofibers. Each muscle cell is packed with bundles of actin and myosin filaments, organized into myofibrils that extend the length of the cell. Packed end to end in a myofibril is a chain of sarcomeres, the functional units of contraction. The internal organization of the filaments gives skeletal muscle cells a striated appearance.
(b) Smooth muscle is composed of loosely organized spindle-shaped cells that contain a single nucleus. Loose bundles of actin and myosin filaments pack the cytoplasm of smooth muscle cells. These bundles are connected to dense bodies in the cytosol and to the membrane at attachment plaques.
Skeletal muscle is made up of bundles of multinucleate muscle cells (myofibers). Each cell contains myofibrils that are composed of repeated units of actin and myosin called sacromeres. This repeated unit of actin and myosin gives the muscle cell a striped or striated appearance. Smooth muscle does not have the same arrangement having only one nucleus and containing loose bundles of actin and myosin.
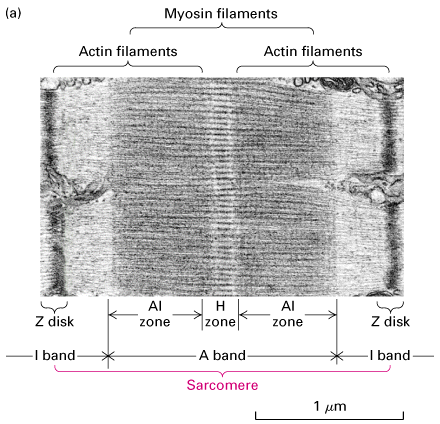
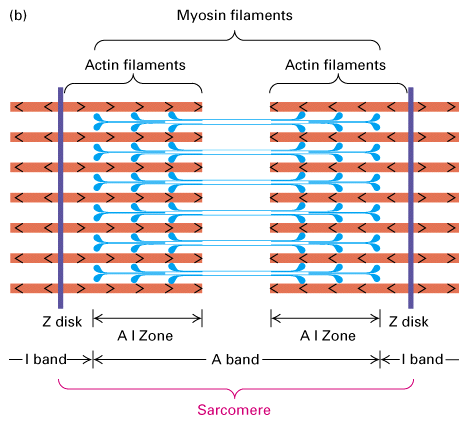
 Figure 18-27,
Lodish 4th edition. Figure 19-22, Lodish 5th edition. Structure of the
sarcomere.
Figure 18-27,
Lodish 4th edition. Figure 19-22, Lodish 5th edition. Structure of the
sarcomere. (a) Electron micrograph of mouse striated muscle in longitudinal section, showing one sarcomere. On either side of the Z disks are the lightly stained I bands, composed of actin filaments. These thin filaments extend from both sides of the Z disk to interdigitate with the dark-stained myosin thick filaments that make up the A band. The region containing both thick and thin filaments (the AI zone) is darker than the area containing only myosin thick filaments (the H zone).
(b) Schematic diagram of a sarcomere. The (+) ends of actin filaments are attached to the Z disks.
(c) Electron micrograph showing actin-myosin cross-bridges in the AI zone of a striated flight muscle of an insect. This image shows a nearly crystalline array of thick myosin and thin actin filaments. The muscle was in the rigor state at preparation. Note that the myosin heads protruding from the thick filaments connect with the actin filaments at regular intervals.
Actin filaments
Figure 18-02, Lodish 4th edition. Structure of F-actin.
Electron micrograph of an actin filament and model of actin. Actin
filaments (F-actin) are made up of individual G-actin molecules that
polymerize into filaments in a process that depends on ATP. The filament is
oriented with a - and a + end, with polymerization occuring faster at the +
end. In places where the cytoskeleton does not change, such as in muscle
sacromeres, the ends of the actin filaments are capped to ensure stability.
(left) Figure 18-03. Binding of myosin S1 head domain to an actin filament.
The myosin heads when bound to actin filaments spiral around the actin.
Myosin thick filament
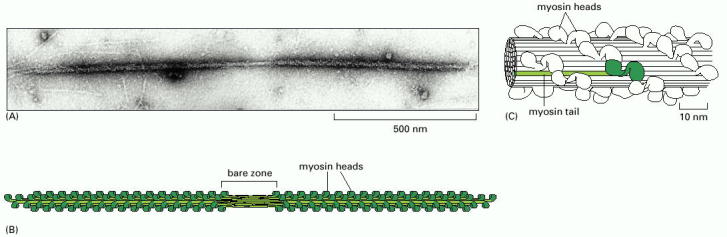
Figure 16-52, Molecular Biology of the Cell, 4th edition. The myosin II bipolar thick filament.
(A) Electron micrograph of a myosin II thick filament isolated from frog muscle. Note the central bare zone, which is free of head domains.
(B) Schematic diagram, not drawn to scale. The myosin II molecules aggregate by means of their tail regions, with their heads projecting to the outside of the filament. The bare zone in the center of the filament consists entirely of myosin II tails.
(C) A small section of a myosin II filament as reconstructed from electron micrographs. An individual myosin molecule is highlighted in green.

Figure 16-70, Molecular Biology of the Cell, 4th edition. Electron micrographs of an insect flight muscle viewed in cross section.
The myosin and actin filaments are packed together with almost crystalline regularity. Unlike their vertebrate counterparts, these myosin filaments have a hollow center, as seen in the enlargement on the right. The geometry of the hexagonal lattice is slightly different in vertebrate muscle.
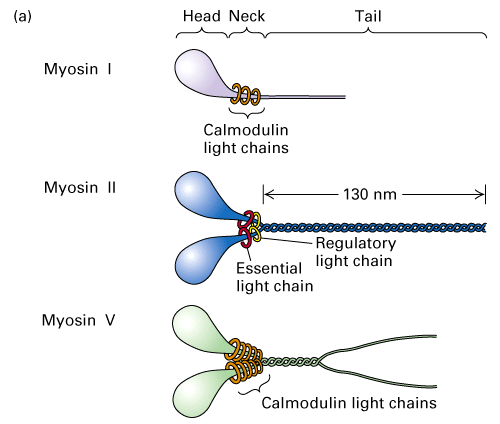
Figure 18-20A, Lodish 4th edition. Structure of some myosin molecules.
There are many members of the myosin family of proteins. Myosin II is
involved in muscle contraction and cytokinesis. The heavy chain of myosin
contains the head domain which is the ATPase domain that binds both actin
and ATP. The neck region behind the head domain binds to the light chains
and the tail domain of two myosin II molecules forms a rodlike structure of
coiled-coiled domains of the myosin II dimer. The light chains associated
with myosin II are Ca+2 binding proteins that help regulate the movement of
myosin along the actin filament.

Figure 18-21B, Lodisth 4th edition. Myosin II coiled tail domains.
The rodlike tail domains of the myosin II dimers associate to form the sacromere thick filament with a bipolar arrangment of head domains at either end.
Sliding filament model
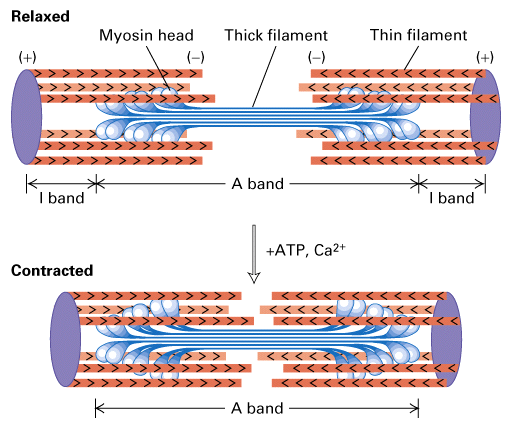
Figure 18-29, Lodish 4th edition. Figure 19-23, Lodish 5th edition. The sliding-filament model of contraction in striated muscle.
The arrangement of thick myosin and thin actin filaments in the relaxed state is shown in the top diagram. In the presence of ATP and Ca2+, the myosin heads extending from the thick filaments pivot, pulling the actin thin filaments toward the center of the sarcomere. Because the thin filaments are anchored at the Z disks (purple), this movement shortens the sarcomere length in the contracted state (bottom).
In the presence of ATP and Ca+2 the myosin heads, extending from the thick filaments, pivot pulling the actin thin filaments towards the center. The thin filaments are anchored and thus the movement shortens the sarcomere length. What would happen if the actin filaments were not anchored?
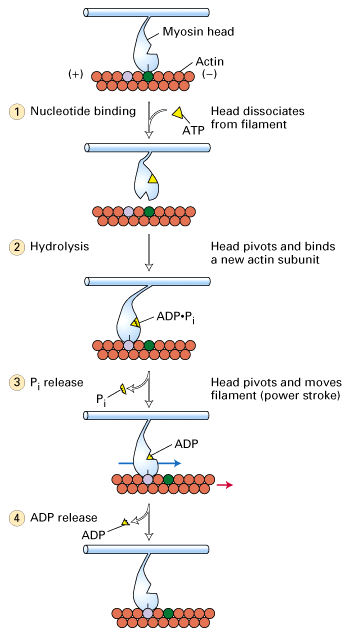
Figure 18-25, Lodish 4th edition. Coupling of ATP hydrolysis to movement of myosin along the actin filament.
A model of how myosin moves is diagrammed in the above figure. It is still not absolutely certain that the myosin head moves once per ATP hyrolyzed but this is the current model. Myosin is bound to actin in the absence of ATP and this is the "rigor" state i.e. gives rigidity to the muscle. ATP binds to the myosin causing the head domain to dissociate from actin. ATP is then hydrolyzed causing a conformational change in the mysoin head to move it to a new position and bind to actin. Pi is released causing the myosin head to change conformation again and it is this movement that moves the actin filament.
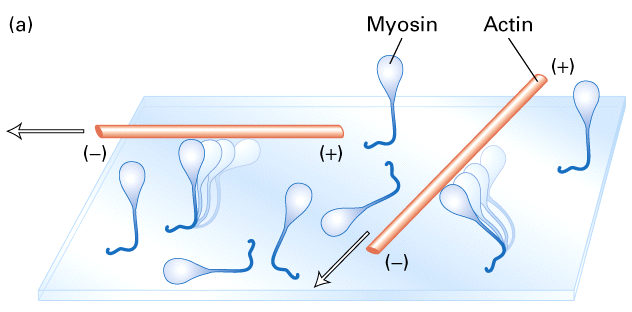
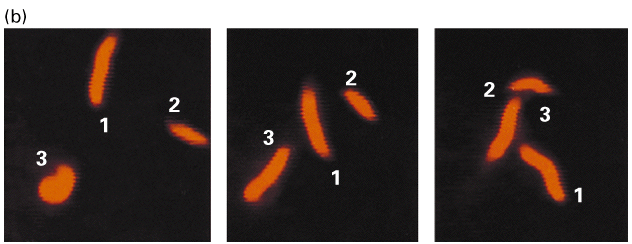
Figure 18-22, Lodish 4th edition. The sliding-filament assay.
Go to this cool web page to see a movie of the movement of actin filaments over myosin. Click here!
Myosin heads slide or walk along the actin filament. Myosin molecules anchored to a glass slide will move actin filaments in the presence of ATP. Figure 18-22 diagrams an experiment where myosin molecules attached to a glass coverslip. Actin filaments that are labelled with a fluorescent dye are then added. In the absence of ATP there is no movement. In the presence of ATP there is movement. Figure B shows the movement of the labelled actin filaments at 30 second intervals. Myosin II from skeletal muscle has the fastest rates of movement. The myosin head moves in discrete steps (5-10 nm in the case of Myosin II) along the actin filament.
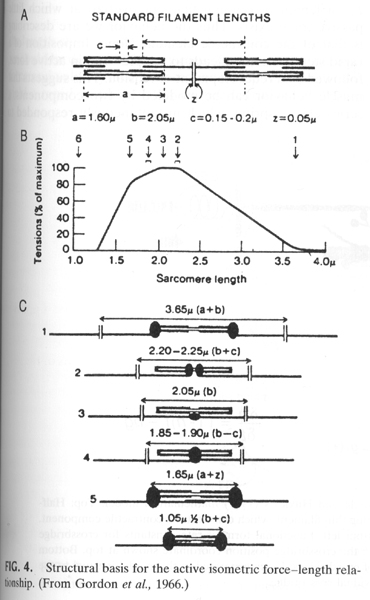
This model predicts that the force of the contraction is proportional to the shortening of the sacromere or the overlap between the thin and thick filaments. Figure 4 below shows this where as the force of the contraction increases the length of the sacromere decreases see B and C.
From "Cell Physiology Source Book"
What keeps the thick and thin filaments linked to the Z line?
Why doesn't the force of myosin against the actin to pull away from the Z
line. I.e. what anchors the filaments to generate the tensions?
The Z disk is complex of proteins that results in the darkly electron dense region. This complex is responsible for anchoring the actin filaments to ensure that the sacromere will shorten during contraction. The actin filaments are capped at both ends to ensure that the actin will not depolymerize. In smooth muscle which do not have Z disks the dense bodies anchor the loose bundles of thin filaments at one end and at the other the actin filaments are anchored in the cell membrane at attachment plaques (similar in many ways to dense bodies).
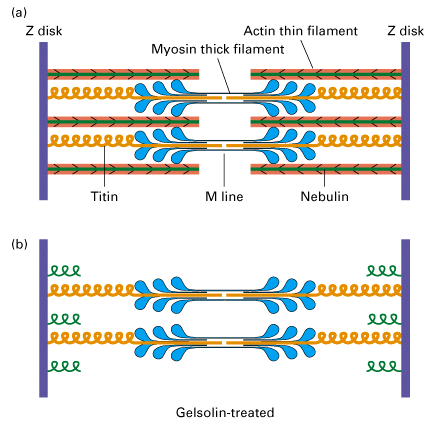
Figure 18-30, Lodish 4th edition. The titin-nebulin filament system stabilizes the alignment of thick and thin filaments in skeletal muscle.
(a) A titin filament attaches at one end to the Z disk and spans the distance to the middle of the thick filament. Thick filaments are thus connected at both ends to Z disks through titin. Nebulin is associated with a thin filament from its (+) end at the Z disk to its (?) end. The large titin and nebulin filaments remain connected to thick and thin filaments during muscle contraction and generate a passive tension when muscle is stretched.
(b) To visualize the titin filaments in a sarcomere, muscle is treated with the actin-severing protein gelsolin, which removes the thin filaments. Without a supporting thin filament, nebulin condenses at the Z disk, leaving titin still attached to the Z disk and thick filament.
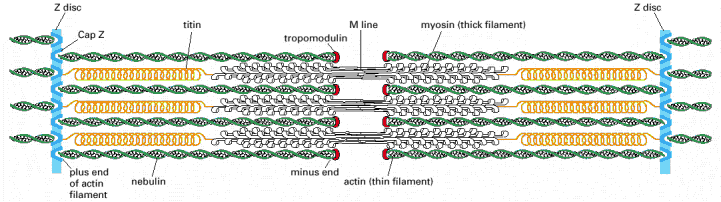
Figure 16-72, Molecular Biology of the Cell, 4th edition. Organization of accessory proteins in a sarcomere.
Each giant titin molecule extends from the Z disc to the M line—a distance of over 1 mm. Part of each titin molecule is closely associated with a myosin thick filament (which switches polarity at the M line); the rest of the titin molecule is elastic and changes length as the sarcomere contracts and relaxes. Each nebulin molecule is exactly the length of a thin filament. The actin filaments are also coated with tropomyosin and troponin (not shown; see Figure 16-74) and are capped at both ends. Tropomodulin caps the minus end of the actin filaments, and CapZ anchors the plus end at the Z disc, which also contains a-actinin.
How does Ca+2 control the interaction between myosin and actin?
As we have seen above Ca+2 is the controlling element for muscle contraction. In the absence of Ca+2 myosin remains bound to actin but can not move. How does Ca+2 control this interaction. Again this process differs in different muscle types.
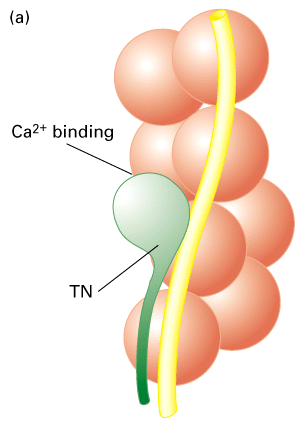
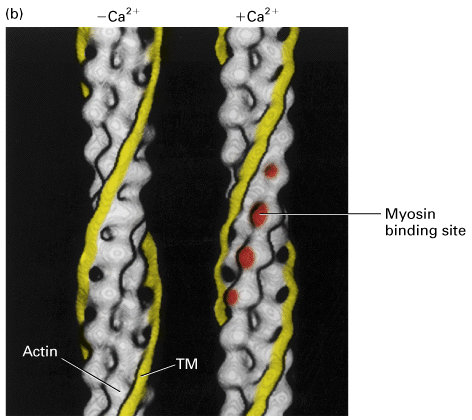
Figure 18-32, Lodish 4th edition. Figure 19-24, Lodish 5th edition. Effect of Ca2+ ions on tropomyosin binding to actin filaments.
(a) Model of the tropomyosin-troponin (TM-TN) regulatory complex on a thin filament. TN, a clublike complex of TN-C, TN-I, and TN-T subunits, is bound to the long a-helical TM molecule.
(b) Three-dimensional electron-microscopic reconstructions of the TM helix (yellow) on a thin filament from scallop muscle. TM in its “off” state (left) shifts to its new position (arrow) in the “on” state (right) when the Ca2+ concentration increases. This movement exposes myosin binding sites (red) on actin. (TN is not shown in this representation.)
In vertebrate skeletal muscle the actin filament is associated with a protein complex that is made up of tropomyosin and troponins C, I and T. Tropomyosin is a ropelike protein that is strung together in a head to tail chain along the actin filament. Tropomyosin contains actin binding sites that ensures a tight interaction. The troponin complex of C, T and I are associated with tropomyosin and controls the location of the tropomyosin on the actin filament. Ca+2 binds to the troponin C subunit and somehow this interaction moves the troponin/tropomyosin complex on the actin filament. This movement now frees up binding sites for myosin II and allows the myosin II to move along the actin filament.
Therefore while there is an increased concentration of Ca+2 then myosin is free to slide along the actin filament. Once the Ca+2 is removed back into the sarcoplasmic reticulum then the troponon/tropomyosin complex moves back and blocks the further movement of myosin along the actin. The sacromere relaxes and the system is set to contract again.
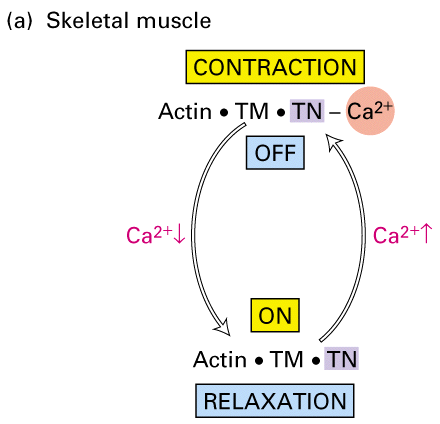
Figure 18-33A. Regulation of skeletal muscle contraction by Ca+2 binding to
troponin.
Skeletal muscle metabolism
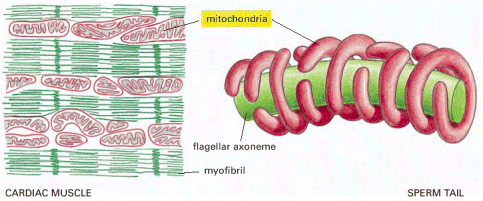
Figure 14-6, Molecular Biology of the Cell. Localization of mitochondria near sites of high ATP utilization in cardiac muscle and a sperm tail.
During the development of the flagellum of the sperm tail, microtubules wind helically around the axoneme, where they are thought to help localize the mitochondria in the tail; these microtubules then disappear, and the mitochondria fuse with one another to create the structure shown.
Muscles of course require a large source of ATP to allow for contraction
and for transport of Ca+2. The ATP requirements are normally met by
glycolysis or respiration. Frequently used high activity muscles use
oxidative respiration mainly to generate ATP. These muscles have extremely
extensive mitochondria which extend through out the myofibrils. They are red
coloured due to a large blood flow and myoglobin (which stores oxygen).
During intense activity though the supply of oxygen can not keep up with the
demand to fuel oxidative phosphorylation even with the reserves from
myoglobin. At this point anaerobic respiration kicks in and glucose is
converted to lactate. This is not very efficient and only generate 2 ATP per
glucose.
Skeletal muscles contain large glycogen stores to help fuel ATP production.
As well skeletal muscles contain creatine phosphate that generates ATP
through :
creatine phosphate + ADP = creatine + ATP
This system actually maintains ATP levels at a similar level in inactive
versus active muscles (this is not the case during intense activity).
The breakdown of glycogen stores in muscles can be stimulated by both Ca+2
and epinephrine. See section on
epinephrine in the cardiac muscle lectures.